
Строение вирусной частицы гепатита в
Текущая версия страницы пока не проверялась опытными участниками и может значительно отличаться от версии, проверенной 31 декабря 2018;
проверки требуют 3 правки.
Ви́рус гепати́та B (англ. Hepatitis B virus, HBV) — ДНК-содержащий вирус из семейства гепаднавирусов, возбудитель вирусного гепатита B. В мире по различным оценкам от 3 до 6 % людей инфицировано вирусом гепатита B. Носительство вируса не обязательно сопровождается гепатитом, однако носитель вируса может заражать других людей.
Классификация[править | править код]
Выделяют 10 генотипов вируса: A, B, C, D, E, F, G, H, I и J. Различия между генотипами составляют более 8 %. Генотипы A и D распространены повсеместно; генотипы C и B характерны для Юго-Восточной Азии и Японии. Генотип Е распространен преимущественно в Африке. Генотип F был найден среди коренного населения Южной Америки и на Аляске. Генотип G спорадически встречается в различных частях света, в частности в Соединенных Штатах Америки и во Франции. Генотипы Е и G характеризуются низкой вариабельностью в последовательности нуклеотидов в геноме, по сравнению с другими генотипами.
Генотипы вируса гепатита B могут обладать различными биологическими свойствами. В последнее время все большое значение придаётся генотипу вируса в клинических аспектах течения вирусной инфекции, а также чувствительности к антивирусным препаратам. На сегодняшний день установлено, что инфекция, вызванная вирусом гепатита B генотипов В и С, коррелирует с повреждением печени; а инфекция, обусловленная вирусом гепатита B генотипа А, эффективно излечивается терапевтическими методами с использованием интерферона
Строение вирусной частицы[править | править код]
Вирион (вирус вне клетки хозяина) состоит из нуклеопротеида, капсида и суперкапсида. Капсид — икосаэдр размером 28 нм, внутри которого заключена ДНК вируса (нуклеопротеид). В состав капсида входит основной белок ядра (HBcAg — от «Hepatitis B core Antigen»). Суперкапсид представляет собой липидную мембрану, в которую встроены молекулы белка наружной оболочки — поверхностного антигена (HBsAg — от «Hepatitis B surface Antigen»). Диаметр вирусной частицы — 42 нм.
Строение генома[править | править код]
Геном вируса гепатита B представляет собой одну двуцепочечную кольцевую ДНК длиной около 3200 нуклеотидов (длина генома различна у разных изолятов). Одна из цепей («плюс-цепь») ДНК короче другой (её длина составляет 1700—2800 нуклеотидов). Вторая цепь тоже не является замкнутой и к её 5’-концу ковалентно присоединена молекула полимеразы (P).
В геноме вируса найдено 4 гена: S, С, Р, X, кодирующих соответственно HBsAg, HBcAg, полимеразу и белок-регулятор экспрессии генов. Кроме того, там есть регуляторные последовательности ДНК, ответственные за синтез белков и репликацию вируса. Гены частично перекрываются, что обусловливает малый размер генома.
Отличительной особенностью генома вируса гепатита В является его высокая информационная ёмкость (особенно на фоне того, что его ДНК является наименьшей по размеру среди всех известных ДНК-содержащих вирусов). Это достигается тем, что в молекуле ДНК, заключающей в себе геном данного вируса, имеется 6 перекрывающихся рамок считывания генов[2].
Репликация вируса[править | править код]
Несмотря на то, что вирус гепатита B является ДНК-содержащим вирусом, в его жизненном цикле имеется РНК-стадия. После проникновения вируса в клетку вирусная ДНК перемещается в клеточное ядро и транскрибируется с образованием полноразмерной прегеномной РНК трёх мРНК меньшей длины. Синтезированные РНК перемещаются в цитоплазму, где мРНК транслируются и прегеномная РНК вместе с полимеразой упаковывается в белковую оболочку. Полимераза вируса гепатита B способна осуществлять обратную транскрипцию, и на матрице прегеномной РНК синтезируется вирусная минус-цепь, а на её матрице — плюс-цепь.
Кроме того, геном вируса полностью или частично может интегрироваться в геном гепатоцита. При этом могут повреждаться гены гепатоцита, что приводит к раку печени.[источник не указан 3440 дней] По другим данным, вирус находится в клетке в эписомном состоянии, а онкогенным эффектом обладает белок Х — транскрипционный фактор, рекрутирующий транскрипционную машину клетки для синтеза РНК вируса[3]
Профилактика[править | править код]
Упомянутый выше поверхностный HBsAg-антиген, находящийся в наружной оболочке, применяют для производства высокоэффективной профилактической вакцины.
Примечания[править | править код]
Источник
Впервые
частицы вируса гепатита В были обнаружены
Д. Дейном в 1970 г. и впоследствии названы
частицами Дейна.
Структура
и химический состав. Вирионы, или частицы
Дейна, имеют сферическую форму диаметром
42 нм. Сердцевина вириона — нуклеокапсид
в форме икосаэдра — состоит из 180
капсомеров. Снаружи он окружен
липосодержащей внешней оболочкой. В
состав вириона входят ДНК, белки,
ферменты, липиды и углеводы.
Строение
вируса гепатита В
Схема
строения вируса гепатита В
Структура
генома HBV необычна. Он состоит из кольцевой
двунитевой молекулы ДНК, которая в
отличие от ДНК других вирусов имеет
однонитевой участок. Его длина непостоянна
и составляет 15—60 % длинной цепи. Кольцевая
молекула ДНК может принимать линейную
форму. В изолированном виде она не
обладает инфекционными свойствами. В
составе вирусного генома обнаружено
около 6 генов, которые контролируют
образование антигенов, структурных
белков и не менее двух ферментов
(ДНК-полимераза, протеинкиназа).
Строение генома вируса гепатита в (strand– нитка)
Антигены. В
составе вируса гепатита В обнаружено
4 антигена: HBs, HBc, НВе и НВх.
HBs-антиген
(ранее именовался австралийским
антигеном) представляет собой гликопротеин
с липидным компонентом, который содержится
во внешней оболочке вириона. В его
составе обнаружено два полипептидных
фрагмента. Один из них (preSg) является
полиглобулиновым рецептором, ответственным
за адсорбцию вируса на аналогичных
рецепторах гепатоцитов. Он связывается
с сывороточным альбумином, который при
полимеризации превращается в полиальбумин.
Таким образом, в составе внешней оболочки
вируса гепатита В имеются те же
полиальбумины, которые содержатся в
сыворотке крови человека. Второй фрагмент
(preSi) обладает выраженными иммуногенными
свойствами. Этот пептид, полученный
генноинженерными методами, может быть
использован для приготовления вакцины.
HBs-антиген обнаруживается в крови.
НВс-антиген
является нуклеопротеином. Он содержится
в сердцевине вирионов, находящихся в
ядрах гепатоцитов, но не поступающих в
кровь.
НВе-антиген
отщепляется от НВс-антигена при его
прохождении через мембрану гепатоцитов,
вследствие чего обнаруживается в крови.
НВх-антиген
наименее изучен. Возможно, он имеет
отношение к раковой трансформации
гепатоцитов.
В
организме больных гепатитом В синтезируются
антитела к трем антигенам HBs, HBc, НВе.
HBsAg =
поверхностный серцевинный антиген
(белок) (4
фенотипа : adw, adr, ayw and ayr)
HBcAg =
внутренний серцевинный антиген (белок)
(единственный
серотип)
HBeAg =
белковый антиген
Репродукция. Вирус
гепатита В не репродуцируется в культурах
клеток и куриных эмбрионах. Репликация
и транскрипция вирусного генома
происходит в ядрах гепатоцитов. При
этом короткая цепь в кольцевой молекуле
ДНК достраивается по длинной цепи с
помощью ДНК-полимеразы, после чего
начинается репликация обеих нитей.
Необычным является возможность
транскрибирования с вирусной ДНК
молекулы РНК, выполняющей функцию
матрицы для синтеза вирусной ДНК путем
обратной транскрипции. Это происходит
при участии имеющегося в зараженных
гепатоцитах фермента обратной
транскриптазы, происхождение которой
неизвестно. Данный процесс, понятный в
случае РНК-содержащих вирусов, которые
с помощью обратной транскрипции получают
возможность встроить свой геном
(ДНК-транскрипт) в хромосому клетки
хозяина, остается загадочным для
ДНК-содержащего вируса гепатита В.
Одновременно
с вирусного генома транслируется
информация для синтеза на рибосомах
гепатоцитов НВс- и HBs-антигенов,
вирусспецифических ферментов и капсидных
белков. Синтезированные нити ДНК
собираются в нуклеокапсиды. При выходе
из клетки они приобретают внешнюю
оболочку с HBs- и НВс-антигенами.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
Источник
Гепатит В –
инфекционное заболевание человека,
характеризуется избирательным поражением
печени и вызывается гепадновирусом
(семейство Hepadnaviridae,
род
Orthohepadnavirus).
Морфология
и структура.
Вирионы гепатита В человека имеют
сферическую форму, диаметр 42 нм, кубический
тип симметрии, состоят из 180 капсомеров.
Геном образует кольцевая двухнитевая
молекула ДНК, состоящая из 3200 нуклеотидов,
но ее «плюс»-нить на 20-50% короче
«минус»-нити. Полный вирион состоит из
внешней липид-гликопротеидной оболочки
(суперкапсид), включающей белковые
молекулы, несущие поверхностный АГ –
HBs АГ
(рис.32).
Суперкапсид HВV
состоит из главного или основного
S-белка, среднего М-белка и большого или
длинного L-белка; имеется внутренний
нуклеокапсид (ядро) диаметром 25-27 нм,
фермент ДНК-полимераза и HBc
АГ, который
содержится в сердцевине вирионов. При
протеолитическом гидролизе белка
капсида образуется полипептид – НВе
АГ – который
отщепляется от НВс АГ при прохождении
его через мембрану гепатоцитов и
обнаруживается в крови.
В
составе поверхностного HBs АГ имеется
один общий АГ а
и две пары взаимоисключающих детерминант
d/у
и w/r.
Известны геномы HВV четырех основных
субтипов, названные по сочетанию АГных
эпитопов HBs Аg: adw,
ayw,
adr,
ayr.
АГы обеспечивают формирование общего
перекрестного иммунитета ко всем
субтипам вируса. Субтипы вируса имеют
различное распространение в регионах
мира. HBs АГ, его полипептидный фрагмент
preS2,
играет важную роль в прикреплении вируса
к гепатоцитам за счет связывания их с
альбуминовым рецептором. Полипептид
preS1
обладает иммуногенными свойствами и
используется для приготовления вакцины.
В зараженной клетке HBs АГ синтезируется
в цитоплазме и участвует в сборке
вирионов. Наряду с полными вирионами
(частицами Дейна) в сыворотке инфицированных
лиц присутствует свободный HBs Аg в виде
сферических частиц диаметром 20-22 нм или
волокнистых образований до 200 нм. Эти
частицы не содержат вирусной ДНК и
являются неинфекционными. Ядерный АГ
HBс АГ в свободной форме локализован в
ядре гепатоцитов и экспрессируется на
поверхности зараженных гепатоцитов,
на которых и может быть обнаружен, но
не выявляется в сыворотке крови больного.
При попадании HBс
АГ в кровь он трансформируется в HBе АГ,
что свидетельствует о высокой инфекционной
опасности больного.
HBх АГ
менее изучен. Считают, что он имеет
отношение к раковой трансформации
гепатоцитов.
Резистентность.Вирус
гепатита В обладает высокой устойчивостью
к температуре, выдерживает кипячение
в течение 15-20 минут, 600С
– несколько часов, в комнатной температуре
сохраняет жизнеспособность в течение
3 месяцев, в замороженном виде – несколько
лет. Вирус погибает при обработке 3%
Н2О2,
5% хлорамином, при УФ-облучении, при
обработке сухим жаром при 180°С в течение
60 минут.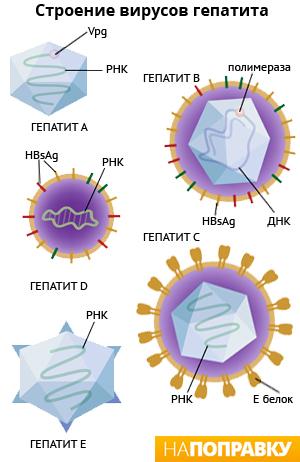
Эпидемиология.
Источником заражения является больной
человек и вирусоноситель. Заражение
может происходит при медицинских
манипуляциях, при переливании крови;
при половом контакте, плод может
заражаться трансплацентарно или через
инфицированные пути матери. Инфицирование
возможно через зубные щетки.
Репродукция.
Вирус гепатита В не репродуцируется в
культурах клеток и куриных эмбрионах.
Репликация происходит в ядрах гепатоцитов.
Она является весьма сложной и состоит
из нескольких этапов. Первоначально
ДНК-полимераза достраивает короткую
(+)цепь ДНК до полной молекулы. Эта ДНК
проникает в ядро клетки, где с нее
синтезируется РНК-копия, получившая
название прегенома.
После этого вирусная ДНК-полимераза
начинает строить на матрице РНК прегенома
его ДНК-копию (обратная транскрипция).
Прегеном одновременно разрушается. Эта
ДНК может встраиваться в геном гепатоцита
(интегративная инфекция). С
вирусного генома транслируется информация
для синтеза на рибосомах гепатоцитов
HBs АГ, HBе АГ, капсидных белков,
вирусспецифических ферментов. Новая
вирусная частица выходит из гепатоцита
(рис. 33). Наряду с вновь образованными
вирусными частицами из инфицированной
клетки высвобождаются также «пустые»
вирусные частицы, содержащие HBs АГ и HBе
АГ. При типичной острой форме гепатита
В в крови появляются HBs АГ, HBе АГ, и АТ
IgM, IgG анти-HBс АГ, анти-HBе АГ и анти-HBs АГ.
Патогенез и
клиника.
Входными путями служат кровеносные
сосуды слизистых оболочек. Вирус попадает
в кровь, заносится в печень и фиксируется
на гепатоцитах. Вирус не обладает
цитопатогенным действием, поэтому
патологический процесс в печени возникает
не с момента внедрения возбудителя в
гепатоциты, а только после распознования
иммуноцитами его АГов на клеточной
мембране, которые индуцируют появление
аутоантител и Т-киллеров к клеткам
печени. Поэтому развивающийся хронический
гепатит и цирроз печени можно рассматривать
как аутоиммунное заболевание.
При развитии
иммунного ответа, представленные на
мембране макрофагов вирусные АГы
индуцируют гуморальный ответ, при
котором образуются АТ IgM и IgG-классов к
HBs АГ, HBс АГ, HBе АГ. Одновременно появляются
АГспецифические Т-киллеры, разрушающие
гепатоциты, на месте которых развивается
соединительная ткань, печень подвергается
склерозу, функция ее нарушается.
Инкубационный
период длится от 30 дней до 6 месяцев.
Болезнь может протекать в латентной
форме, выявляемой только лабораторными
методами, в типичной желтушной форме и
тяжелой («фульминантной») форме,
заканчивающейся летально от печеночной
недостаточности. На фоне увеличения
сывороточных аланиновой и аспарагиновой
аминотрансфераз у 30% взрослых больных
развивается желтуха, которая сохраняется
несколько недель, реже несколько месяцев.
В разгар заболевания и до 8 недель после
него определяются HBs АГ и HBе АГ. Почти
одновременно начинается продукция АТ
к HBс.
Опасность
представляет переход болезни в хроническую
форму, который возможен у 15% взрослых
больных и до 90% новорожденных от больных
матерей. В свою очередь, хронический
В-гепатит является фактором риска
развития цирроза и гепатоцеллюлярной
карциномы.
Лабораторная
диагностика
Для диагностики
применяют определение АГ и АТ в ИФА,
РИА, ПЦР:
—
НВs АГ –
основной маркер острой или хронической
формы инфекции, а также вирусоносительства;
— ДНК НВV –
непосредственный показатель инфицирования
НВV, свидетельствует о репликации вируса;
— Нbе
АГ – маркер острой формы гепатита,
кратковременно выявляемый в крови; его
сохранение более 2 месяцев служит
признаком развития хронического
гепатита;
— IgМ АТ анти-НВс –
главный маркер острой формы заболевания;
— АТ анти-НВs,
анти-НВс, анти-НВе – маркеры завершения
острой формы инфекции, указывающие на
формирование иммунитета к НВV; наличие
только IgG анти-НВs АГ может быть результатом
предшествующей вакцинации или ранее
перенесенной инфекции.
Cпецифическая
профилактика.
Вакцины получены геноинженерным путем,
содержат НВs АГ. По рекомендации ВОЗ
прививки против гепатита В являются
обязательными и должны проводиться
детям на первом году жизни. Используют
генноинженерную вакцину (например,
«Engerix
B»),
в которой использован рекомбинантный
клон дрожжей, содержащий ген НВs+
и вырабатывающий HBs АГ. Полный курс
прививки состоит из 3 инъекций: 1 доза –
сразу после рождения, 2 доза – через 1-2
месяца, 3 доза – до конца 1-го года жизни.
Вакцинировать
необходимо также лиц, которые имеют
повышенный риск инфекции: персонал
медицинских учреждений, больных, которым
проводилось переливание крови и др. Для
экстренной пассивной иммунопрофилактики
используют гамма-глобулин.
Для леченияинтерферон, ингибитор обратной
транскрипции ламивудин, а также
глюкокортикостероиды.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Источник
Структура генома HBV необычна. Он состоит из кольцевой двунитевой молекулы ДНК, которая в отличие от ДНК других вирусов имеет однонитевой участок. Его длина непостоянна и составляет 15—60 % длинной цепи. Кольцевая молекула ДНК может принимать линейную форму. В изолированном виде она не обладает инфекционными свойствами. В составе вирусного генома обнаружено около 6 генов, которые контролируют образование антигенов, структурных белков и не менее двух ферментов (ДНК-полимераза, протеинкиназа).
Строение генома вируса гепатита В
(strand– нитка)
Антигены. В составе вируса гепатита В обнаружено 4 антигена: HBs, HBc, НВе и НВх.
HBs-антиген (ранее именовался австралийским антигеном) представляет собой гликопротеин с липидным компонентом, который содержится во внешней оболочке вириона. В его составе обнаружено два полипептидных фрагмента. Один из них (preSg) является полиглобулиновым рецептором, ответственным за адсорбцию вируса на аналогичных рецепторах гепатоцитов. Он связывается с сывороточным альбумином, который при полимеризации превращается в полиальбумин. Таким образом, в составе внешней оболочки вируса гепатита В имеются те же полиальбумины, которые содержатся в сыворотке крови человека. Второй фрагмент (preSi) обладает выраженными иммуногенными свойствами. Этот пептид, полученный генноинженерными методами, может быть использован для приготовления вакцины. HBs-антиген обнаруживается в крови.
НВс-антиген является нуклеопротеином. Он содержится в сердцевине вирионов, находящихся в ядрах гепатоцитов, но не поступающих в кровь.
НВе-антиген отщепляется от НВс-антигена при его прохождении через мембрану гепатоцитов, вследствие чего обнаруживается в крови.
НВх-антиген наименее изучен. Возможно, он имеет отношение к раковой трансформации гепатоцитов.
В организме больных гепатитом В синтезируются антитела к трем антигенам HBs, HBc, НВе.
HBsAg = поверхностный серцевинный антиген (белок) (4 фенотипа : adw, adr, ayw and ayr)
HBcAg = внутренний серцевинный антиген (белок) (единственный серотип)
HBeAg= белковый антиген
Репродукция. Вирус гепатита В не репродуцируется в культурах клеток и куриных эмбрионах. Репликация и транскрипция вирусного генома происходит в ядрах гепатоцитов. При этом короткая цепь в кольцевой молекуле ДНК достраивается по длинной цепи с помощью ДНК-полимеразы, после чего начинается репликация обеих нитей. Необычным является возможность транскрибирования с вирусной ДНК молекулы РНК, выполняющей функцию матрицы для синтеза вирусной ДНК путем обратной транскрипции. Это происходит при участии имеющегося в зараженных гепатоцитах фермента обратной транскриптазы, происхождение которой неизвестно. Данный процесс, понятный в случае РНК-содержащих вирусов, которые с помощью обратной транскрипции получают возможность встроить свой геном (ДНК-транскрипт) в хромосому клетки хозяина, остается загадочным для ДНК-содержащего вируса гепатита В.
Одновременно с вирусного генома транслируется информация для синтеза на рибосомах гепатоцитов НВс- и HBs-антигенов, вирусспецифических ферментов и капсидных белков. Синтезированные нити ДНК собираются в нуклеокапсиды. При выходе из клетки они приобретают внешнюю оболочку с HBs- и НВс-антигенами.
Репродуктивный цикл HBV
Патогенез заболевания человека и иммунитет. Поскольку входными воротами инфекции при гепатите В являются кровеносные сосуды, вирус сразу же попадает в кровь, с которой разносится по всему организму, фиксируясь прежде всего на гепатоцитах. Однако репродукция вируса не сопровождается цитолизом данных клеток. Это свидетельствует о том, что вирус гепатита В не обладает прямым цитопатическим действием, а патологический процесс в печени возникает не с момента внедрения возбудителя в гепатоциты, а только после распознавания иммуноцитами его антигенов на наружной мембране этих клеток. Таким образом, поражение клеток печени при гепатите В является иммунообусловленным.
Многообразие патогенетических форм данного заболевания (острые, подострые, хронические или персистирующие) определяется антигенами возбудителя и характером его взаимодействия с гепатоцитами, в результате которого развивается продуктивная или интегративная инфекция, с одной стороны, формой иммунного ответа и выраженностью иммунопатологических процессов — с другой.
При острых формах гепатита В подавляется активность Т-хелперов, при хронических — в процесс вовлекаются Т-супрессоры. При стабильном подавлении клонов Т-супрессоров формируются условия для развития аутоиммунных реакций, направленных против собственных клеточных антигенов и прежде всего печеночного липопротеина. При ингибировании Т-хелперов нарушается распознавание вирусных антигенов, что в конечном итоге приводит к подавлению антителообразования.
В то же время HBV наряду с гепатоцитами может взаимодействовать с макрофагами и даже встраивать свою ДНК в их клеточный геном. При -нормальном развитии иммунного ответа представленные на мембране макрофагов вирусные антигены индуцируют гуморальный ответ, который заканчивается синтезом HBs-, HBc- НВе-антител. Это приводит к формированию напряженного гуморального иммунитета, вследствие чего повторные заболевания встречаются редко.
При поражении HBV макрофагов так же, как и Т-хелперов, возникают дефекты в системе распознавания антигенов, что сопровождается развитием иммунодефицитных состояний, являющихся основной причиной персистирующих форм гепатита В.
Интеграция вирусной ДНК в геном гепатоцитов происходит как при острой, так и хронической формах гепатита. При этом она носит случайный характер, так как в каждом случае в хромосому гепатоцитов встраивается неопределенный участок вирусной ДНК- При этом интегрированная вирусная ДНК может оказаться дефектной, что делает невозможным экспрессию ее генов, в том числе и тех, которые контролируют образование антигенов. При интеграции полноценной вирусной ДНК синтезируются вирусные антигены, из которых HBs поступают в кровь. При этом реакции иммунного цитолиза не происходит вследствие отсутствия «мишени» на мембране гепатоцитов для Т-киллеров и NK-клеток. Упомянутые клетки атакуют лишь гепатоциты, несущие НВс-антиген: чем больше данного антигена представлено на их мембране, тем интенсивнее они разрушаются в результате иммунного цитолиза. При этом происходит массовый выход вирусов из разрушенных клеток и генерализация инфекции.
Источник

